The length of time from fertilization until larvae achieve competency remains unknown for most species of octocorals. Once larvae become competent to metamorphose they must locate a suitable environment in which to settle. Successful recruitment depends upon encounter and selection of appropriate habitat, thus an ability to delay metamorphosis in the absence of a suitable environment may enhance the probability of survival by increasing the time available to locate more favorable conditions. The energetic resources available to developing larvae have been shown to influence both competency period and metamorphosis (references in Ben-David-Zaslow and Benayahu 1998). Richmond (1989) proposed that planula with zooxanthellae may benefit from nutritive algal metabolites, allowing them to extend their competency period. Contrary to Richmond ’s prediction, Ben-David-Zaslow and Benayahu (1998) found no significant difference in competency period or larval longevity between several species of zooxanthellate and azooxanthellate soft corals from the Red Sea . This suggests that for lecithotrophic octocoral larvae, nutrient sources within the oocyte are a key important factor controlling larval competence and longevity, and ultimately dispersal capabilities.
The dispersal abilities of octocoral larvae vary widely, especially among alcyonacean-type octocorals. In brooding species with crawling larvae, settlement occurs in the immediate vicinity of the parent colony (e.g. Alcyonium siderium, Sebens 1983; Capnella gaboensis, Farrant 1986) while brooders with swimming, planktonic larvae may disperse more widely (e.g. Anthomastis ritteri, Cordes et al. 2001; Dendronephthya hemprichi, Dahan and Benayahu 1998). Despite their locomotory abilities, most planular larvae of brooding, shallow-water octocorals appear to settle shortly after release (Brazeau and Lasker 1990; Coma et al. 1995; Excoffen et al. 2004). Observations of newly released larvae have shown that they are negatively buoyant and sink rapidly (Benayahu and Loya 1983; Brazeau and Lasker 1990; Gutierrez-Rodriguez and Lasker 2004). Benayahu and Loya (1987) proposed that short-range dispersal of planula is a common trait among coral reef alcyonaceans and may enhance localized recruitment and contribute to the patchy distribution of species. This strategy may be advantageous in space-limited environments (such as coral reefs) because larvae are immediately presented with appropriate habitat type in which to settle (Sebens 1983 in Benayahu and Loya 1987). While most larvae of brooding octocorals exhibit rapid settlement behavior, the planula of the Caribbean gorgonian-type octocoral, Pseudopterogorgia elisabethae, appear to be an exception. Gutierrez-Rodriguez and Lasker (2004) noted from field observations that while settlement of some planula occurred in close proximity to the parent colony (<5 m), most planula remained in the water column (presumably due to turbulence) and were even transported to the water surface, increasing the potential for advection from the local environment.
It is generally accepted that the larvae of broadcast spawning octocorals (i.e. Pennatulaceans and some alcyonaceans) have greater dispersal capabilities than brooded larvae, however data on long-distance transport of octocoral planula is lacking. Dahan and Benayahu (1998) reported that the planular larvae of the broadcast spawning alcyonacean, Dendronephytha hemprichi, swim actively and have a relatively long competency period (65 days). Such larval life features “undoubtedly promote a considerable larval transport in the field.” (Dahan and Benayahu 1998). The cosmopolitan distribution of some deep-water Pennatulids (e.g. Kophobelemnon stelliferum, Rice et al. 1992; U. lindahli, Tyler et al. 1995) also suggests the potential for widespread dispersal of larvae from broadcast spawning. In addition to deep-water Pennatulaceans, many species of deep-water alcyonaceans display widespread distribution, often covering entire ocean basins despite the intermittent occurrence of suitable hard substrate habitats. However, at the present time very little is known about reproductive processes of deep-water octocorals. Thus, factors contributing to the widespread distribution of many species are unresolved.
Numerous environmental features including substrate, light/dark cues, and water motion may influence coral settlement and successful recruitment. Many species of shallow-water octocoral larvae preferentially settle in shaded microhabitats, such as the underside of settlement plates (Alino and Coll 1989; Zeevi Ben-Yosef and Benayahu 1999; Benayahu and Loya 1987; Dahan and Benayahu 1997 and others). This settlement behavior may be an avoidance response to conditions that exist on the upper surface of a plate, such as high light intensity, low tides, competition from filamentous algae, grazing pressure, and sedimentation (Rogers et al. 1984 in Benayahu and Loya 1987; Benayahu and Loya 1984b). Despite potential unfavorable conditions that may exist on exposed surfaces, Alino and Coll (1989) observed reduced survivorship of larvae among the tropical alcyonaceans Lobophytum crassum and Sinularia conferta resident on the undersides of settlement plates compared to the light-exposed surfaces. The general trend toward settlement in shaded microhabitats appears to decrease with increasing depth (Benayahu and Loya 1987), presumably due to both reduced competition with algae and lowered grazing pressure. Thus, it stands to reason that, in deeper waters, substrate and water motion are key factors influencing octocoral planula settlement. In addition to settlement in shaded microhabitats, alcyonacean larvae exhibited a preference for substrates with turf or crustose coralline algae, rough surfaces, and pits in the substrata (Benayahu and Loya 1984b). Water flow may also be an important factor influencing larval settlement. Benayahu and Loya (1987) reported that settlement in Red Sea alcyonacean coral, X. macrospiculata, occurred predominantly along the edges of deployed settlement plates, a pattern which may be related to regions of low flow associated with turbulent eddies created as water travels over the plates (references in Benayahu and Loya 1987).
Once an octocoral planula locates an appropriate location to settle it undergoes metamorphosis to a feeding polyp. This process involves secretion of mucus for temporary attachment to the substrate during settlement (Benayahu and Loya 1983; and others). There are few reports of larval metamorphosis in octocorals. In the alcyonacean soft coral, P. f. fulvum, attachment to the substrate is followed by development into a cone-shaped polyp with 8 tentacular buds. Within a week to ten days, the tentacles elongate and septa develop inside the polyp. In successive weeks tentacles development pinnules and within the next month additional polyps are added and sclerites are present in the polyps (Benayahu and Loya 1983). Similar processes settlement and metamorphosis have been observed in the alcyonacean soft corals Capnella gaboensis (Farrant 1986) and X. macrospiculata (Benayahu and Loya 1984).
A limited number of studies indicate that successful settlement and recruitment into a population occurs at a low rate, at least among shallow-water alcyonacean octocorals (Farrant 1987; Grigg 1977; Lasker et al. 1998). Farrant (1987) reported that for the temperate soft coral, Capnella gaboensis, the first year survival rate of newly settled colonies was 0.26%. Similarly, for the broadcast spawning alcyonacean, Plexaura kuna, Lasker et al. (1998) estimated annual survival of new colonies at 10-6. Using a Poisson probability distribution, Lasker et al. (1998) calculated that in order to produce a successful a recruitment event 95% of the time, that settlement of 700,000 individuals per year would be required. Their calculated mortality rate for P. kuna settlers of 3% per day yields an extremely low probably for successful recruitment, suggesting that this species may rely on certain environmental circumstances that enhance successful recruitment (e.g. favorable substrata, Gotelli 1988; reduced grazing, Yoshioka 1996). Extremely high post-settlement mortality of new recruits implies that successful settlement may be more closely tied to water column and post-settlement survival than to gamete production and fertilization rates (Lasker et al. 1998). Coffroth and Lasker (1998b) proposed that the life history strategy of P. kuna relies on “large and long-lived genets” which may only achieve successful sexual reproduction very infrequently over the course of the colonies’ multi-decadal lifespan. There is evidence that many species, especially those in deep and temperate waters, are long-lived (Andrews et al. 2002; Risk et al. 2002) and therefore may possess similar reproductive life history characteristics.
Table 1
| FAMILY |
SPECIES |
SEX |
REPRODUCTIVE STRATEGY |
Citation |
| Coenothecalia |
Heliopora coerulea |
separate |
surface brooding (6-8 days) below inflated tentacles forming brood-like chamber |
Babcock 1990 |
SOFT CORALS |
|
|
|
|
| Alcyoniidae |
Alcyonium acaule |
- |
brooding |
Hartnoll 1975 |
| Alcyoniidae |
Alcyonium aspiculatum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Alcyonium digitatum |
separate |
broadcast spawning |
Hartnoll 1975 |
| Alcyoniidae |
Alcyonium hibernicum |
separate |
brooding |
Hartnoll 1975 |
| Alcyoniidae |
Alcyonium molle |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Alcyonium palmatum |
- |
brooding |
Hartnoll 1975 |
| Alcyoniidae |
Alcyonium siderium |
separate |
brooding |
Feldman 1970 |
| Alcyoniidae |
Anthomastis ritteri |
separate |
brooding (in siphonozooids) |
Cordes et al. 2001 |
| Alcyoniidae |
Capnella gaboensis |
separate |
external surface brooding |
Farrant 1986 |
| Alcyoniidae |
Cladiella pachyclados |
separate |
broadcast spawning |
Shinkarenko 1981 |
| Alcyoniidae |
Lobophytum compactum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Lobophytum crassum |
separate |
broadcast spawning |
Yamazato et al. 1983 & Uehara et al. 1987,Coll et al. 1995 |
| Alcyoniidae |
Lobophytum hirsutum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Lobophytum microlobatum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Lobophytum pauciflorum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Lobophytum planum |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Lobophytum sarcophoides |
separate |
broadcast spawning |
Dai 1989 |
| Alcyoniidae |
Minabea robusta |
- |
- |
Utinomi and Imahara 1976 |
| Alcyoniidae |
Paraerythropodium fulvum fulvum |
separate |
external surface brooders (entagle eggs in surface mucus) |
Benayahu and Loya 1983 |
| Alcyoniidae |
Sarcophyton crassocaule |
separate |
broadcast spawning |
Dai 1989 |
| Alcyoniidae |
Sarcophyton cf. ehrenbergi |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Sarcophyton glaucum |
separate |
broadcast spawning |
Benayahu and Loya 1986 |
| Alcyoniidae |
Sarcophyton trocheliophorum |
separate |
broadcast spawning |
Shinkarenko 1981 |
| Alcyoniidae |
Sinularia conferta |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Sinularia cruciata |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Sinularia deformis |
separate |
broadcast spawning |
Alino and Coll 1989 |
|
Sinularia dura |
|
|
Pratt 1903 |
|
Sinularia humesi |
separate |
broadcast spawning |
Benayahu et al. 1990 |
| Alcyoniidae |
Sinularia liptoclados |
separate |
broadcast spawning |
Benayahu et al. 1990 |
| Alcyoniidae |
Sinularia lochmodes |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Sinularia mayi |
separate |
broadcast spawning |
Benayahu et al. 1990 |
| Alcyoniidae |
Sinularia polydactyla |
separate |
broadcast spawning |
Alino and Coll 1989 |
| Alcyoniidae |
Sinularia rigida |
separate |
broadcast spawning |
Benayahu |
| Clavulariidae |
Clavularia crassa |
|
surface brooding |
Weinbery 1986 |
| Clavulariidae |
Clavularia hamra |
separate |
external surface brooders (entagle eggs in surface mucus) |
Benayahu 1989 |
| Clavulariidae |
Clavularia inflata |
- |
surface brooding |
Alino and Coll 1989 |
| Clavulariidae |
Pachyclavularia violacea |
- |
surface brooding |
Alino and Coll 1989 |
| Cornulariidae |
Cornularia komaii |
- |
external surface brooding (spawned eggs retained in space made by closed tentacles) |
Suzuki 1971 |
| Cornulariidae |
Cornularia sagamiensis |
- |
external surface brooding (spawned eggs retained in space made by closed tentacles) |
Suzuki 1971 |
| Neptheidae |
Dendronepthya hemprichi |
separate |
broadcast spawning |
Dahan & Benayahu 1997 |
| Neptheidae |
Dendronepthya sinaiensis |
- |
- |
Benayahu 1997 |
| Neptheidae |
Litophyton arboreum |
separate |
brooding |
Benayahu et al. 1990 |
| Xeniidae |
Anthelia formosa |
|
brooding |
Utinomi 1950 |
| Xeniidae |
Anthelia glauca |
separate |
external & internal brooding |
Kruger et al. 1998 |
| Xeniidae |
Efflatounaria sp. |
separate |
external surface brooders |
Dinesen 1985 |
| Xeniidae |
Heteroxenia coheni |
hermaphroditic |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Heteroxenia elizabethae |
hermaphroditic & gonochoristic, dimorphic |
brooding |
Gohar 1940; Shinkarenko 1981 |
| Xeniidae |
Heteroxenia fuscescens |
hermaphrotic, dimorphic |
external & internal brooding |
Benayahu et al. 1989 |
| Xeniidae |
Heteroxenia ghardaqensis |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Sympodium caeruleum |
separate |
internal brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia biseriata |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia blumi |
separate |
brooding |
Gohar 1940 |
| Xeniidae |
Xenia farauensis |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia garciae |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia hicksoni |
separate |
brooding |
Gohar 1940 |
| Xeniidae |
Xenia impulsatilla |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia macrospiculata |
separate |
external brooding |
Achituv et al. 1992 & Benayahu & Loya 1984 |
| Xeniidae |
Xenia membranacea |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia obscuronata |
separate |
brooding |
Benayahu et al. 1989 |
| Xeniidae |
Xenia umbellata |
separate |
internal brooding |
Benayahu et al. 1989 |
GORGONIANS |
|
|
|
|
| Briareidae |
Briarium asbestinum |
separate |
surface brooding (3-5 days) entrapped in mucus sheet |
Brazeau & Lasker 1990 |
| Briareidae |
Briarium stechei |
separate |
surface brooding |
Alino and Coll 1989 |
| Corallidae |
Corallium rubrum |
separate |
brooder |
Vighi 1970 |
| Gorgoniidae |
Eunicella singularis |
- |
brooder |
Weinberg and Weinberg 1979 |
| Gorgoniidae |
Eunicella stricta |
separate |
brooder |
Theodor 1967 |
| Gorgoniidae |
Leptogorgia virgulata |
separate |
broadcast spawning |
Gotelli 1988 |
| Gorgoniidae |
Pseudopterogorgia bipinnata |
separate |
brooder |
Kinzie 1970 |
| Gorgoniidae |
Pseudopterogorgia elisabethae |
separate |
external brooding |
Gutierrez-Rodriguez and Lasker 2004 |
| Isidiidae |
Acanella arbuscula |
separate |
brooder? |
Lawson 1990 |
| Plexauridae |
Eunicea clavigera |
separate |
brooder |
Kinzie 1970 |
| Plexauridae |
Muricea californica |
separate |
brooder |
Grigg 1979 |
| Plexauridae |
Muricea fructicosa |
separate |
brooder |
Grigg 1979 |
| Plexauridae |
Paramuricea clavata |
separate |
surface brooding |
Coma et al. 1995 |
| Plexauridae |
Plexaura flexuosa |
separate |
broadcast spawning |
Beiring and Lasker 2000 |
| Plexauridae |
Plexaura homomalla |
separate |
broadcast spawning |
Martin 1982 |
| Plexauridae |
Plexaura kuna |
separate |
broadcast spawning |
Lasker |
| Plexauridae |
Pseudoplexaura porosa |
separate |
broadcast spawning |
Kapela and Lasker 1989 |
| Primnoidae |
Ainigmaptilon antarcticum |
separate |
? May be brooder (lrg egg size) |
Orejas et al. 2002 |
| Primnoidae |
Fannyella rossii |
- |
brooder |
Orejas et al. 2002 |
| Primnoidae |
Thouarella variabilis |
separate |
internal brooding |
Brito et al. 1995 |
PENNATULACEANS |
|
|
|
|
| Kophobelemnidae |
Kophobelemnon stelliferum |
separate |
probably broadcast spawning |
Rice et al. 1992 |
| Pennatulidae |
Ptilosarcus guerneyi |
separate |
broadcast spawning |
Chia and Crawford 1973 |
| Pennatulidae |
Pennatula aculeata |
separate |
broadcast spawning |
Eckelbarger et al. 1998 |
| Virgularidae |
Virgularia juncea |
- |
- |
Soong 2005 |
| Veretillidae |
Cavernularia obesa |
- |
- |
Mori and Tanase 1971 |
References
Achituv, Y. and Y. Benayahu. 1990. Polyp dimorphism and functional, sequential hermaphroditism in the soft coral Heteroxenia fuscescens (Octocorallia). Marine Ecology Progress Series 64: 263-269.
Achituv, Y., Y. Benayahu, and J. Haniania . 1992. Planulae brooding and acquisition of zooxanthellate in Xenia macrospiculata (Cnidaria: Octocorallia). Helgolander Meeresunters 46: 301-310.
Alino, P.M. and J.C. Coll. 1989. Observations of the synchronized mass spawning and postsettlement activity of octocorals on the Great Barrier Reef , Australia : Biological Aspects. Bulletin of Marine Science 45(3): 697-707.
Andrews, A.H., E.E. Cordes, M.M Mahoney, K. Munk, K.H. Coale, G.M. Cailliet, and J. Heifetz. 2002. Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia 471(1-3): 101-110.
Babcock, R. 1990. Reproduction and development of the blue coral Heliopora coerulea (Alcyonaria: Coenothecalia). Marine Biology 104: 475-481.
Bayer, F.M. 1973. Colonial organization in octocorals. In: Animal colonies: Development and function through time. R.S. Boardman, A.H. Cheetham, and W.A. Oliver (eds.) Dowden, Hutchinson, and Ross, Stroudsburg, PA. Pp 69-93.
Beiring, E.A. and H.R. Lasker. 2000. Egg production by colonies of a gorgonian coral. Marine Ecology Progress Series 196: 169-177.
Ben-David-Zaslow, R. and Y. Benayahu. 1998. Competence and longevity in planulae of several species of soft corals. Marine Ecology Progress Series 163: 235-243.
Benayahu, Y. 1989. Reproductive cycle and developmental processes during embryogenesis of Clavularia hamra (Cnidaria, Octocorallia). Acta Zoologica 70(1): 29-36.
Benayahu, Y. 1991. Reproduction and developmental pathways of Red Sea Xeniidae (Octocorallia, Alcyonacea). Hydrobiologia 216/217: 125-130.
Benayahu Y. and Y. Loya. 1983. Surface brooding in the Red Sea soft coral Parerythropodium fulvum fulvum (Forskaal, 1775). Biological Bulletin 165: 353-369.
Benayahu, Y. and Y. Loya. 1984a. Life history studies on the Red Sea soft coral Xenia macrospiculata Gohar, 1940. I. Annual dynamics of gonadal development. Biological Bulletin 166: 32-43.
Benayahu, Y. and Y. Loya. 1984b. Life history studies on the Red Sea soft coral Xenia macrospiculata Gohar, 1940. II. Planula shedding and post larval development. Biological Bulletin 166: 44-53.
Benayahu, Y. and Y. Loya. 1985. Settlement and recruitment of a soft coral: Why is Xenia macrospiculata a successful colonizer? Bulletin of Marine Science 36(1): 177-188.
Benayahu, Y. and Y. Loya. 1986. Sexual reproduction of a soft coral: Synchronous and brief annual spawning of Sacrophyton glaucum (Quoy and Gaimard, 1833). Biological Bulletin 170: 32-42.
Benayahu, Y. and Y. Loya. 1987. Long-term recruitment of soft-corals (Octocorallia: Alcyonacea) on artificial substrata at Eliat ( Red Sea ). Marine Ecology Progress Series 38: 161-167.
Benayahu, Y. and M.H. Schleyer. 1998. Reproduction in Anthelia glauca (Octocorallia: Xeniidae.) II. Transmission of algal symbionts during planular brooding. Marine Biology 131: 433-442.
Benayahu, Y., Y. Achituv, and T. Berner. 1988. Embryogenesis and acquisition of algal symbiots by planula of Xenia umbellate (Octocorallia: Alcyonacea). Marine Biology 100: 93-101.
Benayahu, Y., T. Berner, and Y. Achituv. 1989. Development of planulae within a mesogleal coat in the soft coral Heteroxenia fuscensens. Marine Biology 100: 203-210.
Bock, S. 1938. The alcyonacean genus Bathyalcyon. Kungl. Svenska Vetenskapsakad. Handl. (3)16(5): 1-54, plates 1-2.
Brazeau, D.A. and H.R. Lasker. 1989. The reproductive cycle and spawning in a Caribbean gorgonian. Biological Bulletin 176(1): 1-7.
Brazeau, D.A. and H.R. Lasker. 1990. Sexual reproduction and external brooding by the Caribbean gorgonian Briareum asbestinum. Marine Biology 104: 465-474.
Brazeau, D.A. and H.R. Lasker. 1992. Reproductive success in the Caribbean octocoral Briareum asbestinum. Marin Biology 114: 157-163.
Brito, T.A.S., P. Tyler, and A. Clarke. 1997. Reproductive biology of the Antarctic octocoral Thouarella variabilis Wright & Studer, 1889. In: Coelenterate Biology: Proceedings of the 6th International Congress of Coelenterate Biology. J.C. den Hartog (ed.). Nationaal Natuurhistorisch Museum , Leiden . Pp. 63-69.
Chia, F.S. and B.J. Crawford. 1973. Some observations on gametogenesis, larval development and substratum selection of the sea pen, Ptilosarcus guerneyi. Marine Biology 23: 73-82.
Coffroth, M.A. and H.R. Lasker. 1998a. Larval paternity and male reproductive success of a broadcast-spawning gorgonian, Plexaura kuna. Marine Biology 131(2): 329-337.
Coffroth, M.A. and H.R. Lasker. 1998b. Population structure of a clonal gorgonian coral: The Interplay between clonal reproduction and disturbance. Evolution 52: 379-393.
Coll, J.C., P.A. Leone, B.F. Bowden, A.R. Carroll, G.M. Koenig, A. Heaton, R. Nys, M. Maida,and P.M. Alino. 1995. Chemical aspects of mass spawning in corals. 2. (-)-Epi-thunbergol, the sperm attractant in the eggs of the soft coral Lobophytum crassum (Cnidaria: Octocorallia). Marine Biology 123: 137-143.
Coll, J.C. and D. Kelman. 1997. Possible locations for sperm chemo-attractants in the eggs of the soft coral Lobophytum crassum. In The Proceedings of the 8th International Coral Reef Symposium 2: 1251-1254.
Coma, R., and H.R. Lasker. 1992. Small-scale heterogeneity of fertilization success in a broadcast spawning octocoral. Journal of Experimental Marine Biology and Ecology 214: 107-120.
Coma, R. and H.R. Lasker. 1997. Small-scale heterogeneity of fertilization success in a broadcast spawning octocoral. Journal of Experimental Marine Biology and Ecology 214: 107-120.
Coma, R., M. Zabala, and J.M. Gili. 1995. Reproduction and cycle of gonadal development in the Mediterranean gorgonian Paramuricea clavata. Marine Ecology Progress Series 117: 173-183.
Coma, R., M. Zabala, and J.M. Gili. 1995. Sexual reproductive effort in the Mediterranean gorgonian Paramuricea clavata. Marine Ecology Progress Series 117: 185-192.
Cordes, E. E., J.W. Nybakken, and G. VanDykhuizen. 2001. Reproduction and growth of,Anthomastis ritteri (Octocorallia: Alcyonacea) from Monterey Bay California , USA . Marine Biology 138: 491-501.
Dahan, M. and Y. Benayahu. 1997. Reproduction of Dendronephthya hemprichi (Cnidaria: Octocorallia): year-round spawning in an azooxanthellate soft coral. Marine Biology 129: 573-579.
Dahan, M. and Y. Benayahu. 1998. Embryogenesis, planulae longevity, and competence in the octocoral Dendronephthya hemprichi. Invertebrate Biology 117(4): 271-280.
Dinesen, Z.D. 1985. Aspects of the life history of a stolon-bearing species Efflatonaria (Octocorallia: Xeniidae). Proceedings of the 5th International Coral Reef Congress 6: 89-94.
Eckelbarger, K.J., P.A. Tyler, and R.W. Langton. 1998. Gonadal morphology and gametogenesis in the sea pen Pennatula aculeate (Anthozoa: Pennatulacea) from the Gulf of Maine . Marine Biology 132: 677-690.
Excoffen, A.C., F.H. Acuna, M.O. Zamponi, and G.N. Genzano. 2004. Reproduction of the temperate octocoral Tripalea clavularia (Octocorallia: Anthothelidae) from sublittoral outcrops off Mar del Plata , Argentina . Journal of the Marine Biological Association of the UK 84(4): 695-699.
Fadlallah, Y.H. and J.S. Pearse. 1982. Sexual reproduction in solitary corals: Overlapping oogeneic and brooding cycles, and benthic planulas in Balanophyllia elegans. Marine Biology 71: 223-231.
Farrant, P.A. 1985. Reproduction in the temperate Australian soft coral Capnella gaboensis. In Proceedings of the 5th International Coral Reef Congress. Gabrie, C. and B. Salvat (eds.), Antenne-Museum-EPHE, Moorea. Vol. 4. pp. 319-324.
Farrant, P.A. 1986. Gonad development and the planula of the temperate Australian soft coral Capnella gaboensis. Marine Biology 92: 381-392.
Farrant, P.A. 1987. Population dynamics of the temperate Australian soft coral Capnella gaboensis. Marine Biology 96: 401-407.
Fautin, D.G. and R.N. Mariscal. 1991. Cnidaria: Anthozoa. In Microscopic anatomy of invertebrates volume 2: Placozoa, Porifera, Cnidaria, Ctenophora. Edited by F.W. Harrison, and J.A. Westfall. Wiley-Liss , New York . Pp. 267-358.
Fitzsimmons-Sosa, K., P. Hallock, J. Wheaton, K.E. Hackett, and M.K. Callahan. 2004. Annual cycles of gonadal development of six common gorgonians from Biscayne National Park , Florida , USA . Caribbean Journal of Science 40(1): 144-150.
Giese, A.C. and H. Kanatani. 1987. Maturation and spawning. In: Reproduction of marine invertebrates. Vol. 9. A.J. Pearse, J.S. Pearse, and V.B. Pearse (eds.) Boxwood Press, Pacific Grove , California . Pp. 251-329.
Gohar, H.A.F. 1940. Studies on the Xeniidae of the Red Sea . Publications of the Marine Biological Station Ghardaqu 2: 27-118.
Gohar, H.A.F. and H.M Roushdy. 1961. On the embryology of the Xeniidae (Alcyonaria) (with notes on the extrusion of larvae). Publications of the Marine Biological Station Ghardaqu ( Red Sea ) 11: 43-70.
Goldberg, W. and R. Hamilton. 1974. The sexual cycle of Plexaura homomalla. In: Prostaglandins from Plexaura homomalla: Ecology, Utilization, and Conservation of a Major Medical Marine Resource. Bayer, F.M. and A.J. Weinheimer, eds., pp. 58-61. University of Miami Press, Coral Gables .
Gotelli , N.J. 1988. Determinants of recruitment, juvenile growth, and spatial distribution of a shallow-water gorgonian. Ecology 69: 157-166.
Grigg, R.W. 1977. Population dynamics of two gorgonian corals. Ecology 58: 278-290.
Gutierrez-Rodriguez,C. and H.R. Lasker. 2004. Reproductive biology, development, and planula behavior in the Caribbean gorgonian Pseudopterogorgia elisabethae. Invertebrate Biology 123(1):54-67.
Harrison , P.L. and C.C. Wallace 1990. Reproduction, dispersal and recruitment of scleractinians corals. In Coral reefs. Z. Dubinsky (ed.). Elsevier Science Publishers, Amsterdam . Pp. 133-207.
Hartnoll, R.G. 1975. The annual cycle of Alcyonium digitatum. Estuarine, Coastal and Shelf Science 3: 71-78.
Harvell, C.D. and R.K. Grosberg. 1988. The timing of sexual maturity in clonal animals. Ecology 69: 1855-1864.
Highsmith, R.C. 1982. Reproduction by fragmentation in corals. Marine Ecology Progress Series 7: 207-226.
Hughes, R.G. 1989. The functional biology of clonal organisms. Chapman and Hall, London .
Jackson , 1985. Distribution and ecology of clonal and aclonal benthic invertebrates. Pages 297-355 in J.B.C. Jackson, L.W. Buss, and R.E. Cook, editors. Population biology and evolution of clonal organisms. Yale University Press, New Haven , Connecticut , USA .
Kapela, W. and H.R. Lasker. 1999. Size-dependant reproduction in the Caribbean gorgonian Pseudoplexaura porosa. Marine Biology 135: 107-114.
Kinize, R.A. III. 1970. The ecology of the gorgonians (Cnidaria: Octocorallia) of Discovery Bay , Jamaica . PhD Thesis, Yale University .
Kruger, A., M.H. Schleyer, and Y. Benayahu. 1998. Reproduction in Anthelia glauca (Octocorallia: Xeniidae). 1. Gametogenesis and larval brooding. Marine Biology 131: 423-432.
Kukenthal, W. 1919. Gorgonaria. Wissenschaftliche Er Gedinsse der Deutschen Tiefsee Expedition 1898-1899. XIII. Band. 2. Tiel 1. Hälfte. Pp. 1-646.
Larkman, A.U. and M.A. Carter. 1984. The apparent absence of a cortical reaction after fertilization in a sea anemone. Tissue and Cell 16: 125-130.
Lasker, H.R. 1983. Vegetitative reproduction in the octocoral Briareum asbestinum (Pallas). Journal of Experimental Marine Biology and Ecology 72: 157-169.
Lasker, H.R. 1984. Asexual reproduction, fragmentation, and skeletal morphology of a plexaurid gorgonian. Marine Ecology Progress Series 19: 261-268.
Lasker, H.R. 1990. Clonal propagation and population dynamics of a gorgonian coral. Ecology 71(4): 1578-1589.
Lasker, H.R. and K. Kim. 1996. Larval development and settlement behavior of the gorgonian coral Plexaura kuna (Lasker, Kiho, and Coffroth). Journal of Experimental Marine Biology and Ecology 207: 161-175.
Lasker, H.R., K. Kim, and M.A. Coffroth. 1996. Reproductive and genetic variation among Caribbean gorgonians: the differentiation of Plexaura kuna, new species. Bulletin of Marine Science 58(1): 277-288.
Lasker, H.R., D.A. Brazeau, J. Claderon, M.A. Coffroth, R. Coma, and K. Kim. 1996. In situ rates of fertilization among broadcast spawning gorgonian corals. Biological Bulletin 190(1): 45-55.
Lasker, H.R., K. Kim, and M.A. Coffroth. 1998. Production, settlement, and survival of plexaurid gorgonian recruits. Marine Ecology Progress Series 162: 111-123.
Lawson, G.S. 1991. Preliminary evidence for seasonal reproduction in the deep-sea gorgonian Acanella arbuscula. Porcupine Newsletter 5(2): 29-35.
Levitan, R.D. 1993. The importance of sperm limitation to the evolution of egg size in marine invertebrates. American Naturalist 141: 517-536.
Levitan, R.D. 1996. Effects of gamete traits of fertilization in the sea and evolution of sexual dimorphism. Nature 382: 153-155.
Levitan, R.D. 2000. Optimal egg size in marine invertebrates: Theory and phylogenic analysis of the critical relationships between egg size and development time in echinoids. American Naturalist 156: 175-192.
Matthews, A. 1917. The development of Alcyonium digitatum with some notes on the early colony formation. Quarterly Journal of the Microscopical Society 62: 43-94.
McFadden, C.S. 1991. A comparative demographic analysis of clonal reproduction in a temperate soft coral. Ecology 72(5): 1849-1866.
Orejas, C., P.J. Lopez-Gonzalez, J.M. Gili, N. Teixido, J. Gutt, W.E. Arntz. 2002. Distribution and reproductive ecology of the Antarctic octocoral Ainigmaptilon antarcticum in the Weddell Sea . Marine Ecology Progress Series 231: 101-114.
Rice, A.L., P.A. Tyler, and G.J.L. Paterson. 1992. The Pennatulid Kophobelemnon stelliferum (Cnidaria: Octocorallia) in the Porcupine Seabight ( North-east Atlantic Ocean ). Journal of the Marine Biological Association of the U.K. 72: 417-434.
Richmond , R.H. 1987. Energetics, competency, and long-distance dispersal of the planula larvae of the coral Pocillopora damicornis. Marine Biology 93: 527-533
Richmond , R.H. 1989. Competency and dispersal potential of planula larvae of a spawning versus a brooding coral. Proceedings of the 6th International Coral Reef Symposium, Townsville , Australia 2: 827-831.
Rinchevich, B. and Y. Loya. 1979. The reproduction of the Red Sea coral Stylophora pistillata. I. Gonads and planulae. Marine Ecology Progress Series 1: 133-144.
Risk, M.J., J.M. Heikoop, M.G. Snow, and R. Beukens. 2002. Lifespans and growth patterns of two deep-sea corals: Primnoa resedaeformis and Desmophyllum cristagalli. Hydrobiologia 471(1-3): 125-131.
Rogers , C.S., H.C. Fitz, M. Gilnack, J. Beets, and J. Hardin. 1984. Scleractinian coral recruitment patterns at Salt River submarine canyon, St. Croix , US Virgin Islands . Coral Reefs 3: 69-76.
Santangelo, G., E. Maggi, L. Bramanti, and L. Bongiorni. 2004. Demography of the over-exploited Mediterranean red coral (Corallium rubrum L. 1758). Scientia Marina 68(1): 199-204.
Satterlie, R.A. and J.E. Case. 1979. Development of bioluminescence and other effector responses in the Pennatulid coelenterate Renilla kollikeri. Biological Bulletin 157: 506-523.
Schafer , W.G. and H. Schmidt. 1980. The anthozoan egg: Differentiation of internal oocytes structure. In: Developmental and Cellular Biology of Coelenterates. P. Tardent, and R. Tardent (eds.) Elsevier/North-Holland Biomedical Press. New York , pp. 47-52.
Schleyer, M.H., A. Kruger, and Y. Benayahu. 1997. Reproductive strategies of South African corals. In: Proceedings of the 6th International Conference on Coelenterate Biology. J.C. den Hartog (ed.) pp. 429-435.
Sebens, K.P. 1983. The larval and juvenile ecology of the temperate octocoral Alcyonium siderium Verrill. 2. Fecundity, survival, and juvenile growth. Journal of Experimental Marine Biology and Ecology 72: 263-285.
Soong, K., and J.C. Lang. 1992. Reproductive integration in reef corals. Biological Bulletin 183: 418-431.
Tyler, P.A., S.K. Bronsdon, C.M. Young, and A.L. Rice. 1995. Ecology and gametogenic biology of the genus Umbellula (Pennatulacea) in the North Atlantic Ocean . Internationale Revue der gesamten Hydrobiologie 80(2): 187-199.
Uehara, T., M. Sato, and K. Yamazato. 1987. General description of developmental stages in a soft coral Lobophytum crassum Marenzeller. Galaxea 6: 185-193.
Utinomi, H. and Y. Imahara. 1976. A new second species of dimorphic alcyonacean octocoral Minabea from the bays of Sagami and Suruga, with the emendation of the generic diagnosis. Publications of the Set Marine Biological Laboratory 23(3/5): pp.207-211.
Walker , T.A., and G.D. Bull. 1983. A newly discovered method of reproduction in gorgonian coral. Marine Ecology Progress Series 12: 137-143.
Wallace, C.C. 1985. Reproduction, recruitment, and fragmentation in nine species of the coral genus Acropora. Marine Biology 88: 217-233.
Yamazato, K., M. Sato, and H. Yamashiro. 1981. Reproductive biology of an alcyonacean coral, Lobophytum crassum Marenzeller. Proceedings of the 4th International Coral Reef Symposium, Manila , Philippeans 2: 671-678.
Yoshioka, P.M. Variable recruitment and its effects on the population and community structure of shallow-water gorgonians. Bulletin of Marine Science. 59(2): 433-443.
Zeevi Ben-Yosef, and Y. Benayahu. 1999. The gorgonian coral Acabaria biserialis: life history of a successful colonizer of artificial substrata. Marine Biology 135: 473-481.
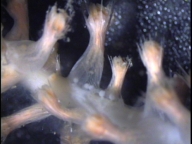
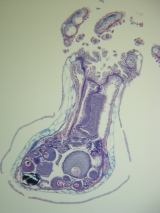 Cross-section of Metallogorgia melanotrichos with developing oocytes.
Cross-section of Metallogorgia melanotrichos with developing oocytes.
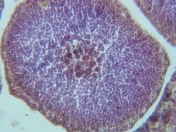 Spermary in Paramuricea placomus from the Gulf of Maine.
Spermary in Paramuricea placomus from the Gulf of Maine.
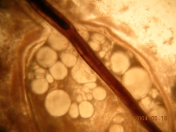 Gonads in Metallogorgia melanotrichos
Gonads in Metallogorgia melanotrichos